



Содержание
Перейти к:
https://doi.org/10.47470/0869-7922-2026-34-3-161-168
EDN: lqfiko
Перейти к:
Введение. Известно, что противоопухолевый препарат доксорубицин (Докс) является репротоксикантом, оказывающим негативное влияние на репродуктивную функцию у млекопитающих. В мужском организме доксорубицин приводит к существенным нарушениям клеток семенников, индуцируя повреждения ДНК, образование двунитевых разрывов, поперечных сшивок, свободных радикалов.
Цель настоящей работы – изучение влияния доксорубицина на развитие клеток сперматогенного эпителия in vitro.
Материал и методы. Исследование проводили на первичной культуре сперматогенного эпителия мыши. Цитотоксическое действие доксорубицина изучали в концентрациях 0,25; 0,05 и 0,005 мкг/мл. Степень цитотоксичности и жизнеспособности клеток оценивали по их морфологическому состоянию, повреждению ДНК, изменению числа живых клеток в культуре в течение 40 суток.
Результаты. Через 24 часа после воздействия Докс отмечали дозозависимое статистически значимое снижение жизнеспособности клеток во всех исследуемых образцах, при этом максимальный эффект – уменьшение численности живых клеток до 50% – отмечен при действии препарата в концентрации 0,25 мкг/мл, уменьшение до 13% – при 0,05 мкг/мл. Снижение жизнеспособности клеток сохранялось на последующих этапах культивирования, что определяло характер монослоя, а также изменения морфологии клеток Сертоли.
Ограничения исследования. Полученные результаты относятся к объекту исследования, который представляет собой гетерогенную клеточную культуру.
Заключение. В ходе экспериментальной работы все исследуемые концентрации доксорубицина оказывали цитотоксическое действие на соматические и сперматогенные клетки. Наиболее губительными для клеток сперматогенного эпителия in vitro были концентрации доксорубицина 0,25 и 0,05, которые приводили к гибели в первые сутки и вызывали повреждения ДНК уже после четырёх часов воздействия. Выявлены характерные особенности патоморфологических изменений в клетках Сертоли. Воздействие Докс даже в малых концентрациях приводит к нарушению формирования фидерного слоя.
Соблюдение этических стандартов. Исследование одобрено комиссией по биоэтике ФГУП «НИИ ГПЭЧ» ФМБА России (протокол № 2 от 28.03.2023 г.).Выведение животных из эксперимента осуществляли согласно ГОСТ 33215–2014 и в соответствии с Европейской конвенцией о защите позвоночных животных, используемых для экспериментов или в иных научных целях (ETS N 123), директивой Европейского парламента и Совета Европейского союза 2010/63/EC от 22.09.2010 г. о защите животных, использующихся для научных целей.
Участие авторов:
Голубенцева Ю.В. – концепция и дизайн исследования, сбор и обработка материала, статистический анализ, написание текста;
Попов В.Б. – написание текста, редактирование;
Радилов А.С. – написание текста, редактирование.
Все соавторы – утверждение окончательного варианта статьи, ответственность за целостность всех её частей.
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов в связи с публикацией данной статьи.
Финансирование. Исследование не имело спонсорской поддержки.
Поступила в редакцию: 28 апреля 2026 / Поступила после исправления: 12 мая 2026 / Принята в печать: 01 июня 2026 / Опубликована: 30 июня 2026
Голубенцева Ю.В., Радилов А.С., Попов В.Б. Цитотоксическое действие доксорубицина на клетки сперматогенного эпителия in vitro. Токсикологический вестник. 2026;34(3):161-168. https://doi.org/10.47470/0869-7922-2026-34-3-161-168. EDN: lqfiko
Golubentseva Yu.V., Popov V.B., Radilov A.S. Cytotoxic effect of doxorubicin on spermatogenic epithelial cells in vitro. Toxicological Review. 2026;34(3):161-168. (In Russ.) https://doi.org/10.47470/0869-7922-2026-34-3-161-168. EDN: lqfiko
Введение
Сперматогенез представляет собой динамичный процесс созревания и развития мужских половых клеток и занимает центральное место в функционировании мужской репродуктивной системы. Одна из распространённых причин мужского бесплодия – патологические изменения и нарушения, индуцированные в различные периоды мужского гаметогенеза химическими, физическими, биологическими факторами. В литературе имеется множество данных о влиянии на мужскую репродуктивную функцию таких химических соединений, как метоксихлор, 2,3,7,8-тетрахлордибензо-диоксин, винклозолин, нитрозометилмочевина, бусульфан, доксорубицин (Докс) и др. [1–4]. Воздействие некоторых токсических веществ во время эмбриогенеза и пренатального периода приводит к многочисленным токсическим эффектам в ходе онтогенетического развития у взрослых животных, проявляющимся атрофией семенных канальцев, снижением численности клеток Сертоли и сперматозоидов, вследствие чего развиваются патологические состояния суб- и инфертильности [2–5]. Показано, что нитрозометилмочевина и дипин приводят к нарушению сперматогенеза у половозрелых мышей при воздействии на самок в период беременности, действуя на незрелые клетки Сертоли и гоноциты, уменьшая пул недифференцированных сперматогониальных стволовых клеток (ССК), долю пахитеновых сперматоцитов, сперматид и приводя к увеличению доли аномальных форм сперматозоидов [3, 6]. Воздействие химических веществ на ранних постнатальных этапах созревания и развития гамет может вызывать нарушения регуляции и поддержания пула ССК семенника, связанные с подавлением экспрессии белков, участвующих в самообновлении и поддержании стволового состояния ССК (ZBTB16 (PLZF), OCT4, NANOG), нарушения процессов дифференциации сперматогоний (SOX3), ингибирование других клеточных процессов. Так, индуцированные нарушения сперматогенеза на этапе созревания дифференцированных сперматогоний могут быть обусловлены нарушением функции белка, кодируемого геном STRA8, необходимого для запуска мейоза и способствующего нормальному созреванию сперматозоидов. У взрослых самцов мышей с мутацией, приводящей к прекращению синтеза транскрипционного фактора, кодируемого Stra8, в гонадах не образуются сперматозоиды, семенные канальцы заполняются сперматогониями типа А, мыши становятся бесплодными [7, 8]. При блокировке или недостаточной функции ядерного фактора – модулятора цАМФ-зависимых чувствительных элементов (CREM) – происходит остановка сперматогенеза на стадии круглых сперматид. Известно, что цитотоксический препарат Докс, используемый при химиотерапии, индуцирует оксидативный стресс в сперматогониях крыс и в клеточных линиях незрелых клеток Сертоли. Препарат приводит к повреждениям ДНК, образованию двунитевых разрывов, аддуктов, поперечных сшивок [9, 10]. S.K. Sah и соавт. показали, что Докс при однократном введении в дозе 10 мг/кг через 14 и 56 суток приводит к уменьшению числа клеток базального слоя сперматогенного эпителия, способствует их вакуолизации, а у взрослых крыс приводит к уменьшению количества сперматозоидов [11].
Поскольку механизмы гаметотоксических эффектов изучены недостаточно, разработка экспериментальных моделей сперматогенеза является актуальной задачей для последующего их использования в токсикологических исследованиях и, что очень важно, развитию исследований по коррекции индуцированного бесплодия. С помощью таких моделей могут быть решены задачи оценки влияния ксенобиотиков на клетки сперматогенного эпителия in vivo и in vitro. Параллельно такие модели позволят разработать условия использования пула донорских ССК, их оптимальный состав для восстановления герминативного эпителия. Для изучения характера повреждений и восстановительного потенциала собственно клеток сперматогенного эпителия значительную ценность приобретают исследования поведения и реакций половых и соматических клеток в экспериментах in vitro с репротоксикантами.
Цель настоящей работы – изучить влияние доксорубицина на развитие клеток сперматогенного эпителия in vitro.
Материал и методы
Характеристика животных. В экспериментах использовали семенники мышей 7–8-го дней после рождения (д.п.р.), полученных от скрещивания половозрелых самок и самцов гибридов CBA×C57Bl/6. Всего в работе были использованы 63 неонатальных самца. Половозрелых самок и самцов мышей-гибридов CBA×С57BL/6F1 получали из Питомника лабораторных животных «Пущино» (г. Пущино, Московская область). Для получения датированной беременности подсаживали двух самок к одному самцу; день обнаружения вагинальной пробки считали первым днём беременности. На 7–8-й дни после рождения проводили эвтаназию неонатальных самцов методом цервикальной дислокации. Содержание и кормление лабораторных животных проводили в соответствии с ГОСТ 33215–2014 «Руководство по содержанию и уходу за лабораторными животными» (2016 г.). Выведение животных из эксперимента осуществляли согласно ГОСТ 33215–2014. Исследование одобрено комиссией по биоэтике ФГУП «НИИ ГПЭЧ» ФМБА России (протокол № 2 от 28.03.2023 г.).
Культивирование in vitro сперматогенных клеток мыши. Выделение клеток семенника проводили в соответствии с методиками [12, 13] в модификации. После эвтаназии животных в стерильных условиях извлекали семенники, помещали в раствор Хэнкса («Биолот», Россия), содержащий 1% смеси антибиотиков (пенициллин 100 ед/мл, стрептомицин 10 мг/мл, «Биолот», Россия), декапсулировали и промывали в фосфатно-солевом буфере («Биолот», Россия). После снятия оболочки семенники нескольких мышей обрабатывали раствором ферментов – 0,7 мг/мл коллагеназы IV типа (Sigma, США), 0,125% трипсин-ЭДТА, 1 мкг/мл ДНКаза (при температуре плюс 37 °C в течение ~10 мин). Затем дозатором механически пипетировали семенники до получения клеточной суспензии, добавляли до 10%-ю фетальную сыворотку плодов коровы (ФСПК), («Биолот», Россия) и центрифугировали (7 мин, 1000 об/мин). Клеточный осадок ресуспендировали дозатором в ростовой среде и высевали в чашки Петри (60 мм), покрытые 0,1%-м раствором желатина (4,5 ∙ 104 кл/см2). В качестве основной ростовой среды использовали DMEM с высоким содержанием глюкозы (4,5 мг/л) («Биолот», Россия) с добавлением 20% ФСПК, 1% смеси антибиотиков («Биолот», Россия), добавки Глутамакс (Gibco, США) и заменимых аминокислот MEM NEAA (Gibco, США) в соответствии с рекомендациями. Для культивирования и дифференцировки сперматогоний использовали такую же ростовую среду, но с пониженным содержанием ФСПК (5%) и добавлением основных ростовых факторов и добавок: Vitamins MEM, EGF, bFGF, GDNF, bovine pituitary extract, тестостерон (Gibco, Sigma, США). Среду заменяли каждые 3–4 дня. Культивирование проводили при температуре плюс 37 °С и газовой фазе 5% СО2 в воздухе.
Для получения постоянных препаратов клетки выращивали на культуральных стёклах, покрытых 0,1%-м раствором желатина, и культивировали в тех же условиях.
Оценка прямого действия доксорубицина на созревание и развитие клеток сперматогенного эпителия in vitro. Выбор изучаемых концентраций обусловлен, прежде всего, соответствием их реальному содержанию в крови животных при однократном введении Докс в дозе 20 мг/кг с учётом экспоненциального убывания концентрации от 0,25 до 0,05 мкг/мл в крови в течение 6–24 ч [14, 15]. Доксорубицин (Sandoz, Словения) добавляли в культуральную среду до конечных концентраций 0,25; 0,05 и 0,005 мкг/мл и инкубировали в течение 24 часов. После воздействия клетки промывали фосфатным буфером и продолжали культивировать описанным выше способом. Забор материала для анализа проводили на 1-е, 6-е, 9-е, 12-е, 16-е, 24-е и 40-е сутки культивирования.
Подсчёт числа живых и погибших клеток в культуре. Количественную оценку клеток в культуре проводили на каждом сроке фиксации в двух повторах с использованием камеры Горяева и светового микроскопа. Жизнеспособность клеток определяли окрашиванием 0,4%-м трипановым синим, анализировали не менее 300 клеток.
Морфологическую оценку живых клеток сперматогенного эпителия мыши проводили с использованием фазово-контрастного микроскопа EclipseTi-S (Nicon, Япония) в определённые исследованием сроки.
Оценку повреждения ДНК (двойные разрывы) в клетках сперматогенного эпителия проводили с использованием антител к биомаркёру γН2АХ через четыре часа после воздействия доксорубицином. Клетки, выращенные на культуральных стёклах, дважды промывали фосфатным буфером и фиксировали в течение 15 минут 4%-м раствором формалина, затем 5 минут в охлаждённом 70%-м этаноле. В качестве положительного контроля на двойные разрывы ДНК клетки культивировали в присутствии перекиси водорода до конечной концентрации 1 мМ при прочих равных условиях. Иммунофлуоресцентный анализ клеток проводили с применением первичных мышиных моноклональных антител против γН2АХ (Millipore, США, 1 : 100; 1,5 ч при температуре плюс 37 °С). В качестве вторых антител использовали конъюгат Goat Anti-mouse IgG Alexa Fluor-488 (Molecular Probes, США, 1 : 500; 1 ч при температуре плюс 37 °С). Промывали препараты раствором PBS в течение 15 минут. Далее препарат монтировали с 10 мкл монтирующей жидкости Vectoshild, содержащей контрастирующий ДНК-связывающий краситель DAPI, для выявления клеточных ядер. Окрашенные препараты анализировали на флуоресцентном микроскопе Zeiss Imager A2 (Германия) с использованием фильтров DAPI, Green, при общем увеличении ×400. В каждом препарате анализировали не менее 300 клеток.
Статистический анализ. Для каждой исследуемой концентрации препарата в ходе эксперимента выполняли не менее трёх независимых повторов. В качестве контроля использовали чашки с клеточной культурой, в которые не добавляли Докс. Статистическую обработку результатов проводили с использованием программного обеспечения GraphPad Prism и MS Excel. С учётом небольшого объёма выборок использовали непараметрические критерии. Для сравнения двух групп использовали U-критерия Манна – Уитни. Данные представлены в виде медианы (Ме) и квартилей (Q1, Q3).
Результаты
Влияние доксорубицина на морфологию и рост клеток сперматогенного эпителия in vitro. Исходное количество клеток до внесения препарата в питательную среду составило 10⁶ кл/мл. Морфологическую оценку клеток в культуре проводили в динамике после воздействия Докс. В контрольных образцах через сутки культивирования наблюдали клетки с чёткими границами, контрастным ядром, гомогенной цитоплазмой (рис. 1, a, см. на вклейке). В ядрах клеток Сертоли визуализировали одно или несколько ядрышек. Половые клетки имели шарообразную форму, крупное ядро, окружённое узким ободком цитоплазмы, высокое ядерно-цитоплазматическое соотношение. Хроматин в ядрах выявлялся. В небольших количествах отмечали наличие других соматических клеток семенника – клеток Лейдига и фибробластоподобных. Соматические клетки демонстрируют высокую способность к адгезии, прикрепляются к пластику и в дальнейшем формируют плотный фидерный слой на 3–4-е сутки культивирования. Через сутки после внесения Докс в культуральную среду наблюдали значительные изменения характера монослоя клеток. Соматические клетки теряли свою адгезивную способность и не прикреплялись к пластику, между клетками образовывались большие пространства. В образцах после воздействия препарата в концентрациях 0,25 и 0,05 мкг/мл наблюдали утрату чётких границ клеток. В цитоплазме клеток Сертоли, фибробластоподобных и половых клетках обнаружили появление вакуолей, клетки становились почти прозрачными (рис. 1, б, см. на вклейке). Наблюдали клетки с неровными границами, в цитоплазме некоторых клеток появлялась зернистость (рис. 1, в, см. на вклейке). Отмечали мутность клеток (нечёткость ядерной картины, размытость). Однако при анализе клеток культуры сперматогенного эпителия после воздействия самой низкой концентрацией (0,005 мкг/мл) в данной серии экспериментов через 24 ч выявляли лишь незначительные морфологические изменения. Почти все клетки имели чёткие границы плазмолеммы и ядра. В единичных клетках Сертоли наблюдали увеличенный объём и зернистость цитоплазмы.
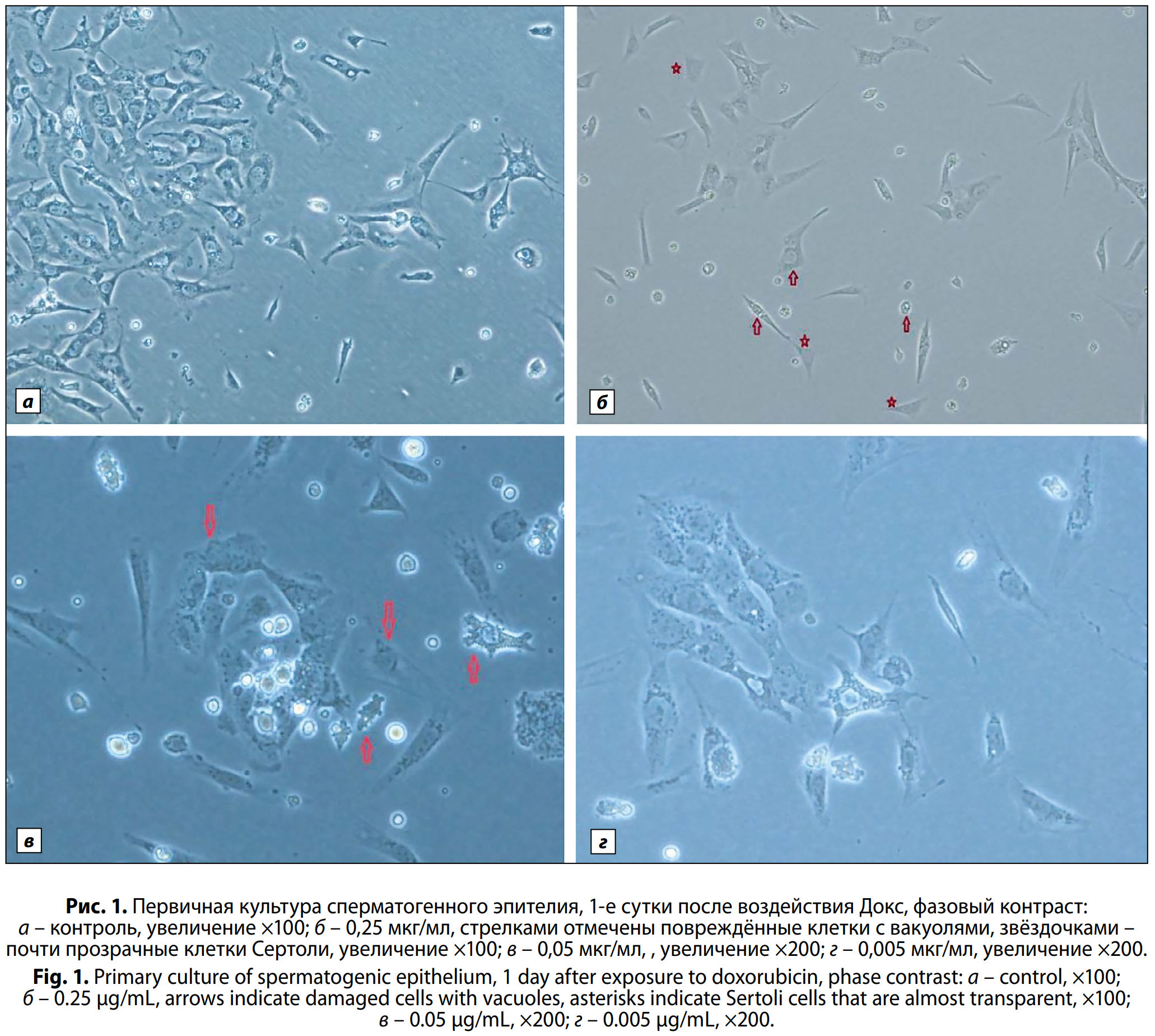
Одним из основных признаков нормального развития и созревания культуры сперматогенного эпителия является формирование плотного фидерного слоя клеток, состоящего преимущественно из клеток Сертоли и малого количества других соматических клеток семенника, на 3–4-е сутки культивирования (с.к.). При анализе образцов клеток на 3–4-е сутки после воздействия (п.в.) ни в одном из экспериментов не образовался нормальный фидерный слой клеток. В образцах после воздействия препарата в концентрациях 0,05 и 0,005 мкг/мл клетки Сертоли лежат неплотно, между ними есть пустые пространства. К 7-м суткам культивирования п.в. препаратом в концентрации 0,005 мкг/мл сформировался фидерный слой, однако в клетках Сертоли наблюдали патологические изменения: множество вакуолей, увеличение объёма цитоплазмы, клетки с остовом цитоплазмы без ядер, со сморщенными ядрами или с увеличенным ядром.
В образцах после воздействия Докс в концентрации 0,25 мкг/мл при наблюдении клеток на 6-е с.к. в фазовом контрасте светового микроскопа отмечали резкое сокращение числа клеток Сертоли. Деструктивные нарушения в клетках продолжали нарастать, и к 9-м суткам культура погибла.
После воздействия препарата в концентрации 0,05 мкг/мл на 4-е и 7-е с.к. также наблюдали аналогичные патологические изменения в клетках Сертоли. Кроме того, наблюдали гигантские клетки Сертоли с большим ядром, исчерченной цитоплазмой, неровными границами и большим количество вакуолей (рис. 2, a, б, см. на вклейке).
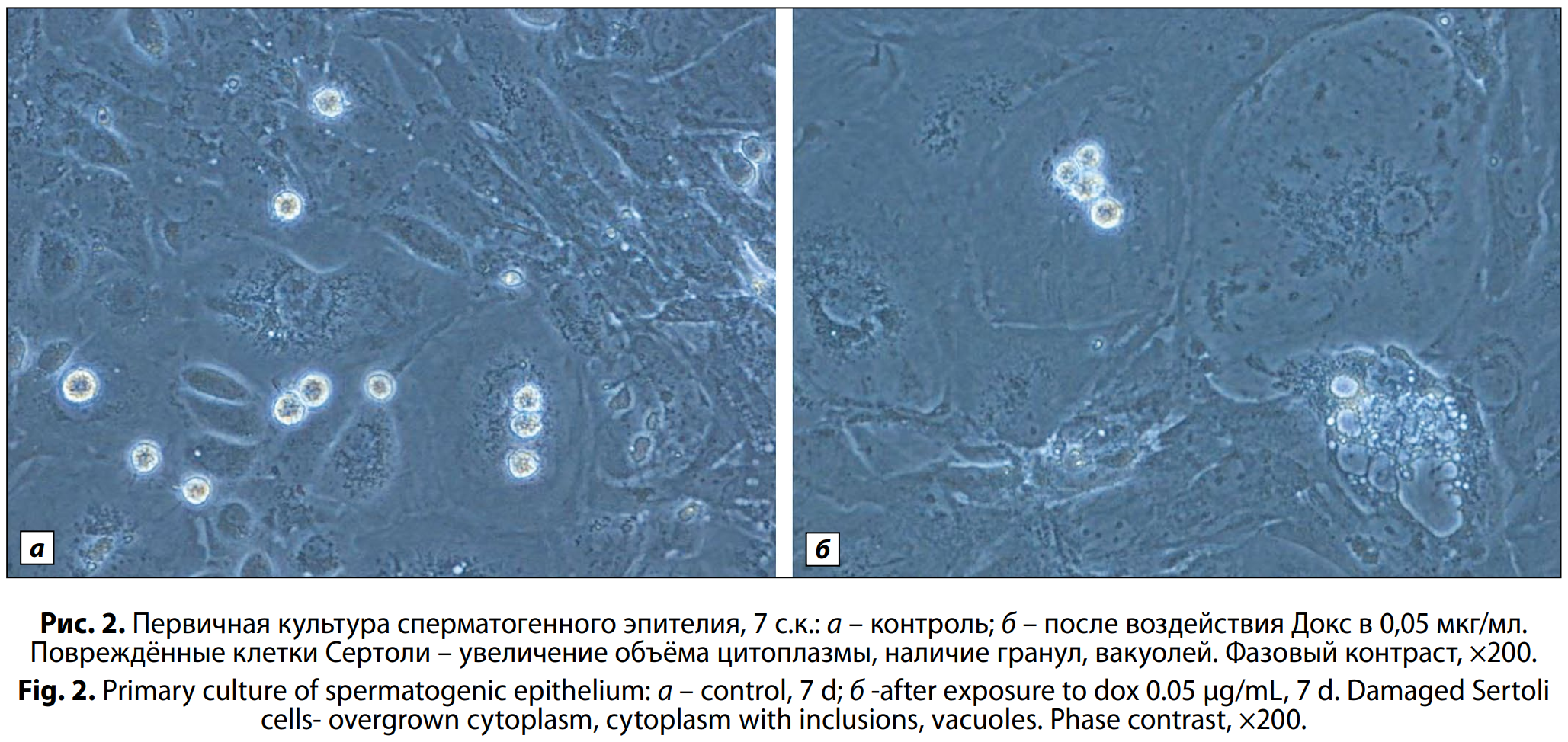
Девятые сутки культивирования при нормальном развитии характеризуются плотным монослоем фидерных клеток и наличием дифференцированных сперматогоний и прелептотенных сперматоцитов, объединённых в небольшие цепочки клеток. В экспериментальных чашках после воздействия Докс в средней и минимальной концентрациях на 9-е с.к. также наблюдаются прелептотенные сперматоциты, но их значительно меньше.
К 16-м суткам культивирования в культуральных чашках без воздействия количество половых клеток увеличивается, отмечается большое количество мейотических клеток, объединённых в группы. После воздействия Докс в минимальной и средней концентрациях на 16-е с.к. наблюдали появление мейотических клеток, круглых сперматид, но в экспериментальных чашках после воздействия Докс в концентрации 0,05 мкг/мл состояние культуры было значительно хуже, сохранялись существенные повреждения клеток Сертоли.
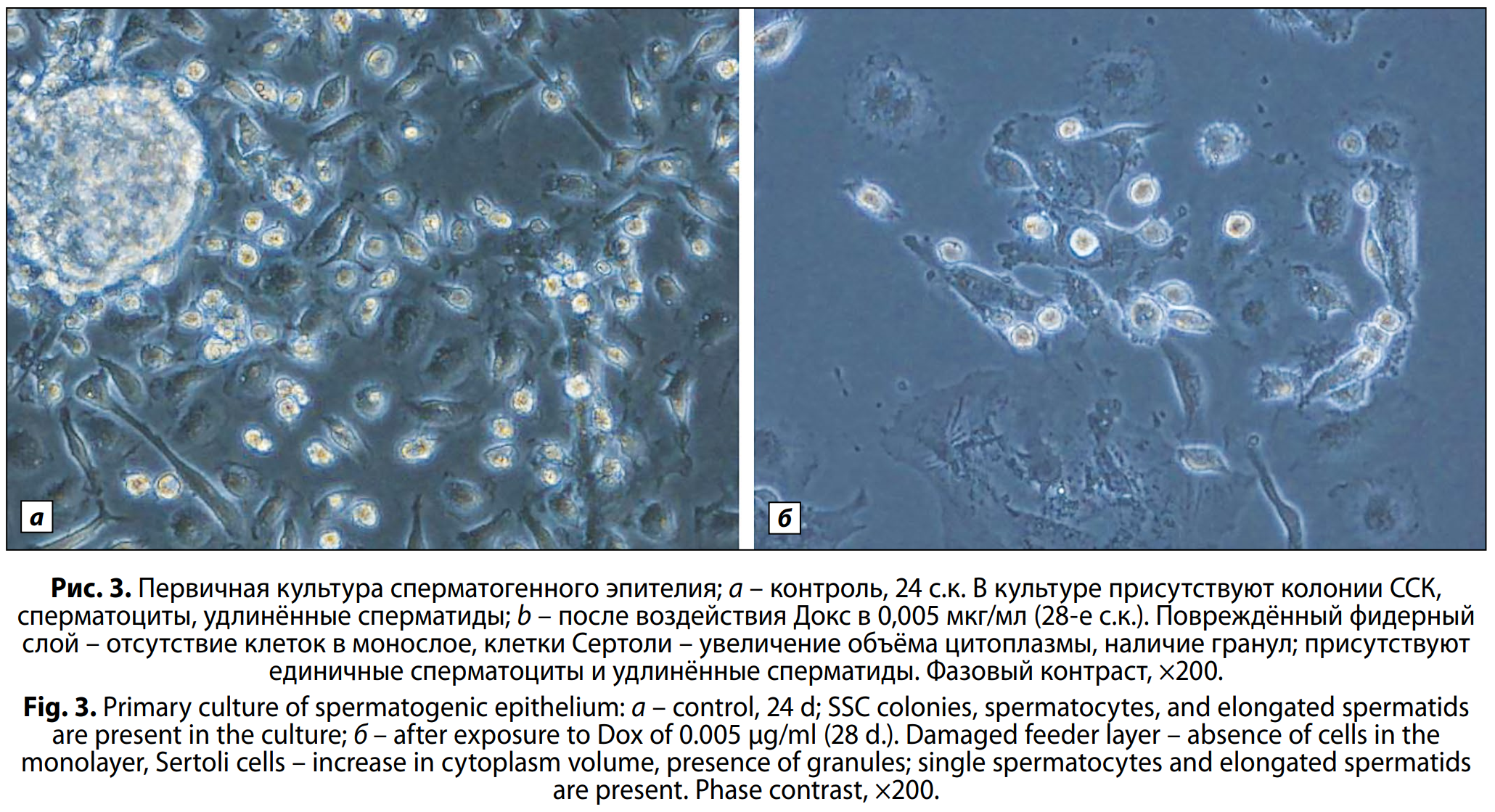
На 24-е с.к. в контроле наблюдали активный сперматогенез с образованием постмейотических клеток – круглых, удлинённых сперматид и сперматозоидов; также присутствовали премейотические и мейотические клетки, объединённые в колонии (рис. 3, a, б, см. на вклейке). В экспериментальных чашках наблюдали наличие круглых, удлинённых сперматид, единичных сперматозоидов, но отмечали отсутствие колоний ССК п.в. Докс в концентрации 0,05 мкг/мл, в то время как при минимальной концентрации ещё присутствовали малые колонии половых клеток. К концу срока наблюдения (40-е с.к.) п.в. Докс в концентрации 0,05 и 0,005 мкг/мл процесс образования сперматид и сперматозоидов затормозился в связи с полным угнетением ССК. В контрольных чашках процесс созревания и развития клеток наблюдали и в последующие дни – до 50–76-х суток культивирования.
Оценка повреждения ДНК клеток с использованием биомаркёра γН2АХ. Двунитевые разрывы ДНК (ДР) часто наблюдаются после воздействия повреждающих факторов и являются реальной угрозой для стабильности генома, в том числе половых клеток. Основная мишень цитотоксических препаратов – пролиферирующие клетки семенника [16–18], однако механизмы, посредством которых происходит развитие тестикулярной токсичности при действии Докс, ещё мало изучены. Появление двунитевых разрывов приводит к фосфорилированию гистона Н2АХ по аминокислотным остаткам Сер¹³⁶ и Сер¹³⁹, образуется γН2АХ – фосфорилированный вариант гистона Н2АХ [19, 20]. Считается, что по уровню фосфорилирования гистона γН2АХ (остатку Сер¹³⁹) можно точно оценить общий уровень повреждения ДНК в клетках [21, 22].
Иммунофлуоресцентный анализ для определения двойных разрывов ДНК выявил, что через 4 часа п.в. Докс доля повреждённых ядер увеличивается в зависимости от концентрации препарата (см. таблицу).

Оценка прямого действия доксорубицина на клетки сперматогенного эпителия in vitro. Для оценки цитоксичности препарата применяли метод, основанный на подсчёте количества клеток в монослое [23]. Полученные данные представлены на рис. 4. При оценке цитотоксичности Докс выявили дозозависимое статистически значимое снижение общего количества клеток во все исследуемые сроки. Через сутки культивирования в присутствии Докс 0,25 и 0,05 мкг/мл отмечали статистически значимое снижение количества клеток – на 50% и 13% соответственно (p = 0,0286, p = 0,0079 соответственно).
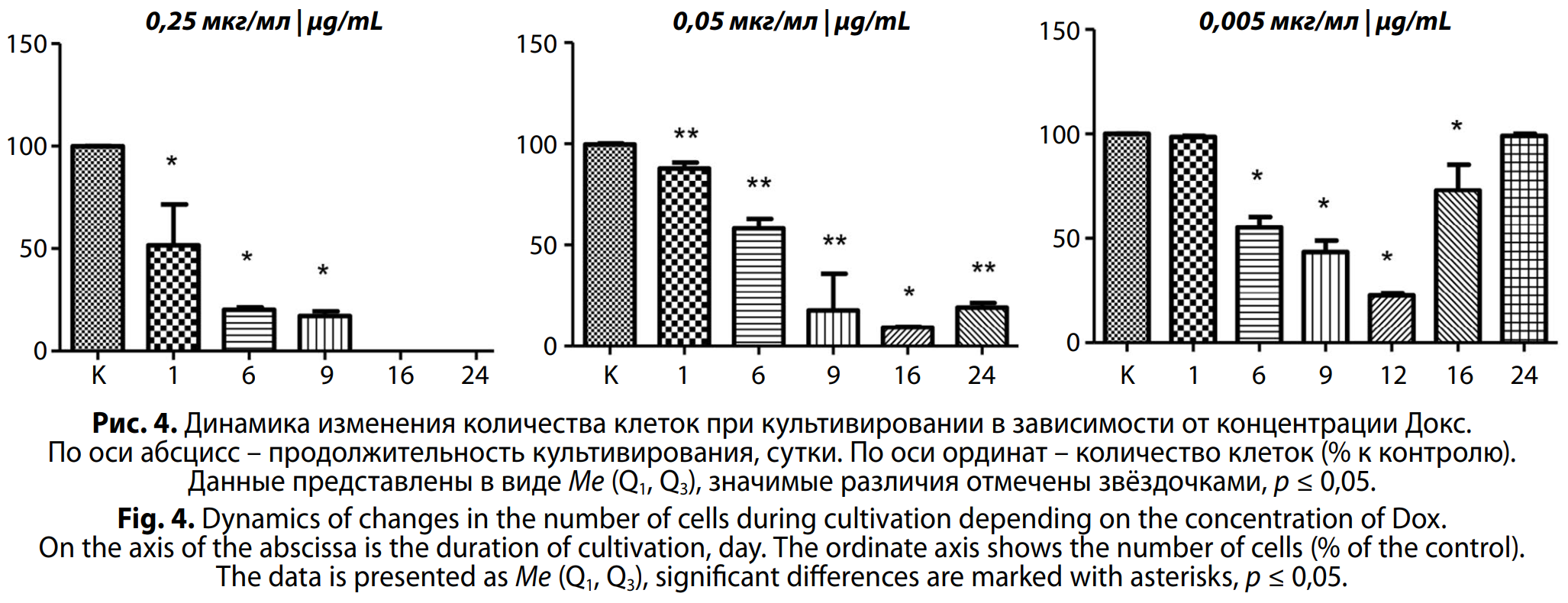
В последующие сроки культивирования наблюдали отдалённый цитотоксический эффект препарата. Так на 6–9-е сутки культивирования гибель клеток возрастала при всех исследуемых концентрациях. В образцах при воздействии Докс 0,25 мкг/мл к 9-м суткам общее количество клеток статистически значимо уменьшилось на 87% (p = 0,0286) (см. рис. 4).
Одновременно проводили оценку жизнеспособности клеток на каждом экспериментальном сроке с помощью окрашивания витальным красителем трипановым синим. Во всех экспериментах, независимо от концентрации Докс, выявлено снижение жизнеспособности клеток через сутки после воздействия. При максимальной концентрации наблюдали гибель клеток до 9-х суток. После воздействия Докс в концентрациях 0,05 и 0,005 мкг/мл общее число жизнеспособных клеток начинало увеличиваться после 12-х суток. (данные не приведены). Это было связано, прежде всего, с началом мейотического процесса. К 16-м суткам наблюдали появление сперматоцитов. Однако, как видно на рис. 4, в образцах клеток при воздействии Докс в концентрации 0,05 мкг/мл процесс восстановления шёл значительно хуже: на 24-е сутки гибель клеток в культуре достигала 81%, в то время как при концентрации 0,005 мкг/мл этот показатель был менее 1%.
Обсуждение
Доксорубицин вызывает дозозависимый эффект при непосредственном воздействии на клетки сперматогенного эпителия в культуре. По морфологическим признакам установлено, что повреждения происходят как в сперматогенных, так и в соматических клетках. Выявлены характерные особенности патоморфологических изменений в клетках Сертоли. Воздействие Докс даже в малых концентрациях приводит к нарушению формирования фидерного слоя, который образуется позднее, чем в контроле. Доксорубицин в концентрации 0,005 мкг/мл демонстрирует отдалённые цитотоксические эффекты. При таком воздействии сохранившиеся сперматогонии, по-видимому, вошедшие в дифференцировку, оказались неспособными к её завершению и формированию сперматозоидов. Возможно также, что воздействие Докс вызвало утрату способности клеток к самообновлению, что неизбежно должно было привести к невозможности запуска новой волны созревания ССК. Таким образом, на клетках сперматогенного эпителия in vitro показаны дозозависимые эффекты Докс как для ССК, так и для соматических клеток.
С учётом особенностей созревания и развития гетерогенной популяции клеток сперматогенного эпителия оценка воздействия доксорубицина должна проводиться практически в течение всего времени наблюдения с учётом отдалённых последствий цитотоксического воздействия, которые реализуются у последующих поколений клеток. Подобные изменения при воздействии на животных (человека) способны приводить к бесплодию. Проблемы и способы коррекции патологических изменений с целью восстановления фертильности в настоящее время изучаются нами и будут представлены в следующих публикациях.
Ограничения исследования. Полученные результаты относятся к объекту исследования, который представляет собой гетерогенную клеточную культуру.
Заключение
В ходе экспериментальной работы все исследуемые концентрации доксорубицина (0,25; 0,05 и 0,005 мкг/мл) проявили цитотоксический эффект на культуре клеток сперматогенного эпителия. Наиболее губительными для клеток сперматогенного эпителия in vitro были концентрации доксорубицина 0,25 и 0,05 мкг/мл, которые приводили к гибели клеток в первые сутки и вызывали повреждения ДНК уже через четыре часа после воздействия. Воздействие Докс на культуру сперматогенного эпителия также вызывает дозозависимые повреждения клеток Сертоли, выражающиеся в их патологических изменениях. В результате наблюдается также зависимое от дозы снижение жизнеспособных клеток Сертоли.
1. Liu S., Wei B., Wei H., Xu A., Sheng L., Sun X., et al. L-cysteine mitigates busulfan-induced testicular injury through modulation of CBS/H2S axis. Front. Cell Dev. Biol. 2025; 13: 1679330. https://doi.org/10.3389/fcell.2025.1679330 https://elibrary.ru/atvfas
2. Hall A., Mattison D., Singh N., Chatzistamou I., Zhang J., Nagarkatti M., et al. Effect of TCDD exposure in adult female and male mice on the expression of miRNA in the ovaries and testes and associated reproductive functions. Front. Toxicol. 2023; 5: 1268293. https://doi.org/10.3389/ftox.2023.1268293 https://elibrary.ru/edpndv
3. Захидов С.Т., Паранюшкина Л.П., Махран Хода Х.А., Эль-Саед К.А.-Х, Голиченков В.А. Влияние химических мутагенов на сперматогенез млекопитающих. Количественная оценка. Известия Российской академии наук. Серия Биологическая. 1994; (6): 870–9.
4. Tahir A., Ijaz M.U., Naz H., Afsar T., Almajwal A., Amor H., et al. Protective effect of didymin against 2, 3, 7, 8-tetrachlorodibenzo-p-dioxin-induced reproductive toxicity in male rats. Naunyn Schmiedebergs Arch. Pharmacol. 2024; 397(4): 2203–14. https://doi.org/10.1007/s00210-023-02763-4 https://elibrary.ru/pubzzr
5. Martini M., Froment P., Franceschini I., Pillon D., Guibert E., Cahier C., et al. Perinatal exposure to methoxychlor affects reproductive function and sexual behavior in mice. Front. Endocrinol. (Lausanne). 2020; 11: 639. https://doi.org/10.3389/fendo.2020.00639 https://elibrary.ru/pfzisk
6. Кулибин А.Ю., Захидов С.Т., Маршак Т.Л., Челомбитько О.М. Ответ сперматогенной системы ускоренно стареющих мышей линии SAMP1 на действие химического мутагена дипина. Известия Российской академии наук. Серия Биологическая. 2008; (3): 272–82. https://elibrary.ru/ijuxgx
7. Mark M., Jacobs H., Oulad-Abdelghani M., Dennefeld C., Féret B., Vernet N., et al. STRA8-deficient spermatocytes initiate, but fail to complete, meiosis and undergo premature chromosome condensation. J. Cell Sci. 2008; 121(Pt. 19): 3233–42. https://doi.org/10.1242/jcs.035071
8. Ma H.T., Niu C.M., Xia J., Shen X.Y., Xia M.M., Hu Y.Q., et al. Stimulated by retinoic acid gene 8 (Stra8) plays important roles in many stages of spermatogenesis. Asian J. Androl. 2018; 20(5): 479–87. https://doi.org/10.4103/aja.aja_26_18 https://elibrary.ru/ippbyx
9. Skinner R., Mulder R.L., Kremer L.C., Hudson M.M., Constine L.S., Bardi E., et al. Recommendations for gonadotoxicity surveillance in male childhood, adolescent, and young adult cancer survivors: a report from the international late effects of childhood cancer guideline harmonization group in collaboration with the PanCareSurFup consortium. Lancet Oncol. 2017; 18(2): e75–90. https://doi.org/10.1016/s1470-2045(17)30026-8
10. Yang F., Teves S.S., Kemp C.J., Henikoff S. Doxorubicin, DNA torsion, and chromatin dynamics. Biochim. Biophys. Acta. 2014; 1845(1): 84–9. https://doi.org/10.1016/j.bbcan.2013.12.002 https://elibrary.ru/yeahxb
11. Sah S.K, Khatiwada S., Chaudhary D., Jha Ch.B., Bhattacharya S. Doxorubicin induced histomorphometric changes in testes of albino rats. Nepal J. Biotechnol. 2015; 3(1): 10–4. https://doi.org/10.3126/njb.v3i1.14223
12. Савченкова И.П., Васильева С.А. Культивирование сперматогоний хряка на клетках Сертоли. Цитология. 2016; 58(2): 135–42. https://elibrary.ru/vkcfqp
13. Волкова Н.А., Коржикова С.В., Котова Т.О., Ветох А.Н., Волкова Л.А., Зиновьева Н.А. Выделение, культивирование и характеристика сперматогониев петуха (Gallus gallus). Сельскохозяйственная биология. 2016; 51(4): 450–8. https://doi.org/10.15389/agrobiology.2016.4.450rus https://elibrary.ru/wkcfqt
14. Drugbank. Doxorubicin. Available at: https://go.drugbank.com/drugs/DB00997
15. Rahman A., Ganjei A., Neffe J.R. Comparative immunotoxicity of free doxorubicin and doxorubicin encapsulated in cardiolipin liposomes. Cancer Chemother. Pharmacol. 1986; 16(1): 28–34. https://doi.org/10.1007/bf00255282
16. Silva R.C., Britto D.M.C., de Fátima Pereira W., Brito-Melo G.E.A., Machado C.T., Pedreira M.M. Effect of short- and medium-term toxicity of doxorubicin on spermatogenesis in adult Wistar rats. Reprod. Biol. 2018; 18(2): 169–76. https://doi.org/10.1016/j.repbio.2018.03.002
17. Habas K., Anderson D., Brinkworth M.H. Germ cell responses to doxorubicin exposure in vitro. Toxicol. Lett. 2017; 265: 70–6. https://doi.org/10.1016/j.toxlet.2016.11.016
18. Brilhante O., Okada F.K., Sasso-Cerri E., Stumpp T., Miraglia S.M. Late morfofunctional alterations of the Sertoli cell caused by doxorubicin administered to prepubertal rats. Reprod. Biol. Endocrinol. 2012; 10: 79. https://elibrary.ru/mragtd
19. Hoeijmakers J.H.J. DNA repair mechanisms. Maturitas. 2001; 38(1): 17–22. https://doi.org/10.1016/S0378-5122(00)00188-2
20. Литвинов С.В. Основные пути репарации двойных разрывов геномной ДНК и взаимодействие между ними. Цитология и генетика. 2014; 48(3): 64–77.
21. Bächler S. Detection of γH2AX foci in animal tissue. Report; 2012. Available at: https://static1.squarespace.com/static/559921a3e4b02c1d7480f8f4/t/598062f09f745638bf643db5/1501586162696/Baechler.pdf
22. Hamer G., Roepers-Gajadien H.L., van Duyn-Goedhart A., Gademan I.S., Kal H.B., van Buul P.P., et al. DNA double-strand breaks and ɤH2AX signaling in the testis. Biol. Reprod. 2003; 68(2): 628–34. https://doi.org/10.1095/biolreprod.102.008672
23. Иксанова А.Г., Бондарь О.В., Балакин К.В. Методы исследования цитотоксичности при скрининге лекарственных препаратов. Казань; 2016.
Научный сотрудник, ФГУП «НИИ гигиены, профпатологии и экологии человека» ФМБА России, 188663, Ленинградская область, Всеволожский район, г.п. Кузьмоловский, Россия
e-mail: yul9olub@yandex.ru
Доктор медицинских наук, профессор, ФГУП «НИИ гигиены, профпатологии и экологии человека» ФМБА России, 188663, Ленинградская область, Всеволожский район, г.п. Кузьмоловский, Россия
e-mail: a.radilov@icloud.com
Доктор биологических наук, ведущий научный сотрудник, ФГУП «НИИ гигиены, профпатологии и экологии человека» ФМБА России, 188663, Ленинградская область, Всеволожский район, г.п. Кузьмоловский, Россия
e-mail: vpopovlr@mail.ru
Голубенцева Ю.В., Радилов А.С., Попов В.Б. Цитотоксическое действие доксорубицина на клетки сперматогенного эпителия in vitro. Токсикологический вестник. 2026;34(3):161-168. https://doi.org/10.47470/0869-7922-2026-34-3-161-168. EDN: lqfiko
Golubentseva Yu.V., Popov V.B., Radilov A.S. Cytotoxic effect of doxorubicin on spermatogenic epithelial cells in vitro. Toxicological Review. 2026;34(3):161-168. (In Russ.) https://doi.org/10.47470/0869-7922-2026-34-3-161-168. EDN: lqfiko
121087, г. Москва, Багратионовский проезд, д. 8, кор. 2.
Филиал РПОХБВ ФБУН «ФНЦГ им. Ф.Ф. Эрисмана» Роспотребнадзора
тел.: +7 (499) 145-60-23
Эл.почта: director@rosreg.info
Открытый доступ

































